Zdieľam:



Typy bunkových zväzkov a ich charakteristiky
bunkové spojenia sú to kontaktné mostíky, ktoré existujú medzi cytoplazmatickými membránami medzi susednými bunkami alebo medzi bunkou a matricou. Kĺby závisia od typu študovaného tkaniva, pričom sa zdôrazňujú súvislosti medzi epiteliálnymi, svalovými a nervovými bunkami.
V bunkách sú molekuly súvisiace s adhéziou medzi nimi. Sú však potrebné ďalšie prvky, ktoré zvyšujú stabilitu väzby tkanív. To sa dosahuje s bunkovými spojeniami.
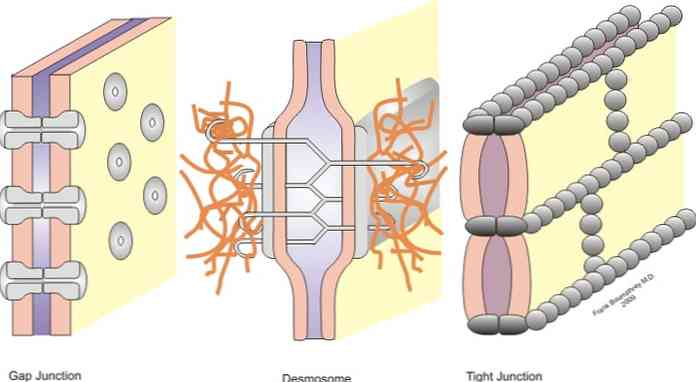
Spoje sú rozdelené do symetrických kĺbov (úzke spoje, desmozómy v páse a štrbinové spoje) a asymetrické spoje (hemidesmosómy).
Úzke križovatky, pásové desmozómy, bodové desmozómy a hemidesmozómy sú kĺby, ktoré umožňujú ukotvenie; keďže spojenia v štrbine sa chovajú ako mosty spojenia medzi susednými bunkami, čo umožňuje výmenu rozpustených látok medzi cytoplazmami.
K pohybu rozpustených látok, vody a iónov dochádza medzi jednotlivými bunkovými zložkami a medzi nimi. Existuje teda transcelulárna dráha, ktorá je riadená radom potrubí a dopravníkov. Na rozdiel od paracelulárnej dráhy, ktorá je regulovaná kontaktmi medzi bunkami - to znamená bunkovými spojeniami.
V rastlinách nájdeme bunkové spojenia, ktoré sa podobajú štrbinovým spojom, nazývaným plazmidmy. Hoci sa líšia svojou štruktúrou, funkcia je rovnaká.
Z lekárskeho hľadiska majú niektoré nedostatky v bunkových spojeniach za následok získané alebo dedičné ochorenia spôsobené poškodením epiteliálnej bariéry..
index
- 1 Charakteristiky
- 2 Typy
- 2.1 - Úzke spojenia
- 2.2 - Slitky alebo medzery
- 2.3 - Kotvenie alebo prípojné spoje
- 2,4-Hemidesmosomas
- 3 Bunkové spojenia v rastlinách
- 4 Lekárska perspektíva
- 5 Referencie
rysy
Živé organizmy sa skladajú z oddelených a rôznorodých štruktúr nazývaných bunky. Tie sú ohraničené plazmatickou membránou, ktorá ich udržuje oddelené od extracelulárneho prostredia.
Napriek tomu, že sú súčasťou živých bytostí, nepodobajú sa tehlám, pretože nie sú od seba izolované.
Bunky sú elementy, ktoré sú navzájom komunikované, ako aj s extracelulárnym prostredím. Preto musí existovať spôsob, akým môžu bunky tvoriť tkanivá a komunikovať, zatiaľ čo membrána zostáva neporušená.
Tento problém je riešený vďaka prítomnosti bunkových spojov, ktoré existujú v epiteli. Tieto spojenia sú vytvorené medzi dvomi susednými bunkami sú klasifikované podľa funkcie každého v symetrických a asymetrických spojoch.
Asymetrické spojenia patria k hemidesmozómom a pri symetrických spojeniach úzke spojenia, desmosóm na páse, desmozómy a štrbinové spoje. Ďalej podrobne opíšeme jednotlivé odbory.
typ
-Úzke križovatky

Úzke križovatky, tiež známe v literatúre ako okluzívne spojenia, sú sektory v bunkových membránach susedných buniek, ktoré sú úzko spojené - ako názov „úzke spojenie“ označuje.
Za priemerných podmienok sú bunky oddelené vzdialenosťou 10 až 20 nm. Avšak v prípade úzkych križovatiek je táto vzdialenosť významne zmenšená a membrány oboch buniek sa dotýkajú alebo dokonca spájajú.
Typická úzka križovatka je umiestnená medzi bočnými stenami susedných buniek v minimálnej vzdialenosti od ich apikálnych povrchov.
V epitelovom tkanive všetky bunky vytvárajú spoje tohto typu, aby zostali spojené. V tejto interakcii sú bunky umiestnené tak, že tvoria vzor pripomínajúci kruh. Tieto odbory pokrývajú celý obvod.
Proteíny zapojené do tesných spojov
Ocludina a Claudina
Oblasti blízkeho kontaktu obklopujú celý povrch bunky. Tieto oblasti tvoria anastomotizujúce kontaktné prúžky transmembránových proteínov známych ako occludin a claudin. Termín anastomóza odkazuje na spojenie určitých anatomických prvkov.
Tieto dva proteíny patria do skupiny tetraespanínov. Sú charakterizované tým, že majú štyri transmembránové domény, dve vonkajšie slučky a dva relatívne krátke cytoplazmatické chvosty.
Bolo pozorované, že occludin interaguje so štyrmi ďalšími proteínovými molekulami, nazývanými occludin zonula a skrátene ZO. Táto posledná skupina zahŕňa proteíny Z01, Z02, ZO3 a afuna.
Claudin je na druhej strane rodina 16 proteínov, ktoré tvoria sériu lineárnych fibríl v úzkych spojeniach, čo umožňuje, aby táto únia prevzala úlohu "bariéry" v paracelulárnej dráhe..
Nectinas a JAM
Nektíny a adhézne molekuly odborov (skrátene skratka v anglickom JAM) sa objavujú aj v úzkych križovatkách. Tieto dve molekuly sa nachádzajú ako homodiméry vo vnútrobunkovom priestore.
Nektíny sú spojené s aktínovými vláknami pomocou afadínového proteínu. Posledne uvedené sa javia ako nevyhnutné, pretože pri deléciách génu kódujúceho afadín u hlodavcov vedú k smrti embrya..
Funkcie úzkych križovatiek
Tieto typy spojov medzi bunkami vykonávajú dve základné funkcie. Prvým je stanovenie polarity buniek v epiteli, oddelenie apikálnej domény od bazolaterálu a zabránenie neprimeranej difúzii lipidov, proteínov a iných biomolekúl..
Ako sme uviedli v definícii, bunky epitelu sú zoskupené v kruhu. Táto štruktúra oddeľuje apikálny povrch bunky od laterálnych a bazálnych, čo vytvára diferenciáciu medzi doménami.
Táto separácia je považovaná za jednu z najdôležitejších koncepcií pri štúdiu fyziológie epitelu.
Po druhé, tesné spojenia bránia voľnému prechodu látok cez vrstvu epitelových buniek, čo sa premieta do bariéry paracelulárnej dráhy..
-Slitky alebo medzery

Slitky alebo medzery sa nachádzajú v oblastiach bez obmedzenia cytoplazmatickej membrány medzi susednými bunkami. V rozdelenom uzle sú spojené cytoplazmy buniek a vytvára sa fyzické spojenie, kde môže nastať prechod malých molekúl..
Táto trieda kĺbov sa nachádza prakticky vo všetkých epiteloch av iných typoch tkanív, kde slúžia na rôzne účely..
Napríklad v niekoľkých tkanivách sa štrbinové štrbiny môžu otvoriť alebo uzavrieť v reakcii na extracelulárne signály, ako je to v prípade neurotransmitera dopamínu. Prítomnosť tejto molekuly znižuje komunikáciu medzi triednymi neurónmi v sietnici v reakcii na zvýšenú intenzitu svetla.
Proteíny podieľajúce sa na štiepení kĺbov
Slitové spojenia sú tvorené proteínmi nazývanými konexíny. Takže "conexón" sa získa spojením šiestich konexínových monomenos. Táto štruktúra je dutý valec, ktorý prechádza cez cytoplazmatickú membránu.
Konexóny sú usporiadané tak, že medzi cytoplazmami susedných buniek je vytvorené potrubie. Okrem toho majú konexóny tendenciu agregovať a tvoriť druh platní.
Funkcie štrbinových spojov
Vďaka tvorbe týchto väzieb sa môže vyskytnúť pohyb určitých molekúl medzi susednými bunkami. Veľkosť molekuly, ktorá sa má transportovať, je rozhodujúca, optimálny priemer je 1,2, ako sú ióny vápnika a cyklický adenozínmonofosfát..
Konkrétne ide o anorganické ióny a molekuly rozpustné vo vode, ktoré môžu byť prenesené z bunkovej cytoplazmy do súvislej cytoplazmy..
V tomto kanáli zohrávajú rozhodujúcu úlohu koncentrácie vápnika. Keď sa koncentrácia vápnika zvyšuje, axiálne kanály majú tendenciu sa uzatvárať.
Týmto spôsobom sa štrbinové spoje aktívne zúčastňujú na procese elektrickej a chemickej väzby medzi bunkami, ako je tomu v bunkách srdcového svalu, ktoré sú zodpovedné za prenos elektrických impulzov..
-Kotviace alebo adherentné spojenia

Pod úzkymi spojmi nájdeme kotviace spoje. Všeobecne sa nachádzajú v blízkosti apikálneho povrchu epitelu. V tejto skupine môžeme rozlíšiť tri hlavné skupiny, zonula adherens alebo desmosoma v páse, makulové adherens alebo desmosome point a desmosome.
V tomto type spojov sú susedné bunkové membrány, ktoré sú spojené zonulami a adhéznymi makulami, oddelené bunkovou vzdialenosťou, ktorá je relatívne široká - ak ich porovnáme s minimálnym priestorom, ktorý existuje v prípade úzkych spojení..
Medzibunkový priestor je obsadený proteínmi, ktoré patria do rodiny kadherínov, desmogleínov a desmokolínov spojených s cytoplazmatickými platňami, ktoré majú iné proteíny nazývané desmoplaquina, placoglobina a plakofilina..
Klasifikácia kotvových spojov
Zonula adherens
Rovnako ako v prípade úzkych spojov v kotviacich spojoch pozorujeme aj usporiadanie usporiadania vo forme krúžku alebo pásu. Zonula adherens je spojená s aktínovým mikrofilmom, prostredníctvom interakcie dvoch proteínov: kadherínov a katenínov..
Macula adherens
V niektorých prípadoch je táto štruktúra známa jednoducho ako desmozóm, je to bodové spojenie, ktoré je spojené s intermediárnymi filamentmi tvorenými keratínom. V tejto súvislosti sa uvedené keratínové štruktúry nazývajú "tonofilimanetos". V epitelových bunkách sa vlákna rozprestierajú od jedného bodu k druhému.
Bod desmosomas
Tieto poskytujú epitelové bunky pevnosť a tuhosť. Predpokladá sa teda, že jeho hlavná funkcia súvisí s posilňovaním a stabilizáciou susedných buniek.
Desmozómy môžu byť porovnané s druhmi nitov alebo zvarov, pretože sa podobajú samostatným drobným škvrnám a nie spojitým pásom.
Zistili sme, tento typ spojov interkalovaných disky, ktoré sa viažu kardiomyocytov v srdcovom svale a mozgových blán obloženie vonkajšieho povrchu mozgu a miechy.
-hemidesmozom

Hemidesmozómy patria do kategórie asymetrických spojov. Tieto štruktúry majú funkciu ukotvenia bazálnej domény epiteliálnej bunky základnou bazálnou laminou.
Termín hemidesmozóm sa používa preto, lebo táto štruktúra sa javí doslova "stredne" desmosóm. Z hľadiska ich biochemického zloženia sú však obe odbory úplne odlišné.
Je dôležité objasniť, že desmozómy sú zodpovedné za priľnutie susednej bunky za inú, zatiaľ čo funkciou hemidesmozómu je pripojiť sa k bunke s bazálnou laminou..
Na rozdiel od makulových adhézií alebo desmozómov majú hemidesmosómy odlišnú štruktúru, ktorá sa skladá z: cytoplazmatickej laminy spojenej s intermediárnymi filamentami a platne externých membrán, ktorá je zodpovedná za spojenie hemidesmozómu s bazálnou laminou, kotevné vlákno.
Jednou funkciou je hemidesmozom zvyšuje celkovú stabilitu epiteliálnych tkanív, a to vďaka prítomnosti cytoskeletálnych medziľahlých vlákien pripojené k zložiek bazálnej membráne.
Bunkové spojenia v rastlinách
Rastlinnej ríši chýba väčšina bunkových spojov opísaných vyššie, s výnimkou funkčného náprotivky, ktorý sa podobá štrbinovým spojom.
V rastlinách sú cytoplazmy susedných buniek spojené dráhami alebo kanálmi nazývanými plazmidmy.
Táto štruktúra vytvára kontinuitu z jednej rastlinnej bunky do druhej. Hoci sa štrukturálne líšia od štrbinových spojov, majú veľmi podobné úlohy, čo umožňuje prechod malých iónov a molekúl.
Lekárska perspektíva
Z hľadiska medicíny sú relevantnými problémami bunkové zväzky. Bolo zistené, že mutácie v génoch, ktoré kódujú proteíny zahrnuté v spojeniach, sú preložené do klinických patológií.
Napríklad, ak je nejaký mutácie v géne pre konkrétny typ Claudin (jeden z proteínov, ktoré sprostredkovávajú interakcie odbory úzky typ) spôsobuje zriedkavé ochorenie v humánnej.
Toto je syndróm straty horčíka v obličkách a symptómy zahŕňajú nízke množstvo horčíka a záchvatov.
Okrem toho sa zistilo, že mutácia v géne, ktorý kóduje proteín nektín 1, je zodpovedná za syndróm rozštepu podnebia alebo rázštepu rtu. Tento stav je považovaný za jednu z najčastejších malformácií u novorodencov.
Mutácie v géne nektínu 1 sú tiež spojené s ďalším stavom nazývaným ektodermálna dysplázia, ktorá postihuje kožu, vlasy, nechty a zuby ľudí..
Pemphigus foliaceus je bulózne kožné patológie určená autoprotilátkami proti desmogleinu 1, prvok kľúč, ktorý je zodpovedný za udržiavanie súdržnosť epidermis.
referencie
- Alberts, B., Bray, D., Hopkin, K., Johnson, A.D., Lewis, J., Raff, M., ... & Walter, P. (2015). Základná bunková biológia. Garland Science.
- Cooper, G.M., & Hausman, R.E. (2000). Bunka: Molekulárny prístup. Sinauer Associates.
- Curtis, H., & Barnes, N. S. (1994). Pozvánka na biológiu. Macmillan.
- Hill, R.W., Wyse, G.A., Anderson, M., Anderson, M. (2004). Fyziológia zvierat. Sinauer Associates.
- Karp, G. (2009). Bunková a molekulárna biológia: koncepty a experimenty. John Wiley & Sons.
- Kierszenbaum, A., & Tres, L. (2016). Histológia a bunková biológia: úvod do patológie. Elsevier Brazília.
- Lodish, H., Berk, A., Darnell, J.E., Kaiser, C.A., Krieger, M., Scott, M.P., ... & Matsudaira, P. (2008). Molekulárna bunková biológia. Macmillan.
- Voet, D., & Voet, J. G. (2006). biochémie. Panamericana Medical.